


CCR’s Earth Fest Events
On April 22, during 12-1PM at Memorial Union, CCR will host a discussion and book signing event, featuring author Jeff Goodell and Professor Andrea Dutton of the Department of Geoscience. Goodell is the author of the New York Times bestseller, The Heat Will Kill You First: Life and Death on a Scorched Planet. The book details the dangers to the environment from rising global temps associated with climate change, particularly the devastating effects on human health.
On April 26, during 2-3:30PM at Union South, CCR will host the following panel discussion: Supporting the Vitality of Rural Communities Through the Wisconsin Rural Partnerships Institute. The panel will focus on the Wisconsin Rural Partnerships Institute (RPI), which supports the state’s rural communities. Panels will include Steve Vavrus (Director, Wisconsin State Climatology Office), Glenda Gillaspy (Dean and Director, College of Agricultural and Life Sciences), Patrick Robinson (Associate Dean, Agriculture, Natural Resources, and Community Development, Division of Extension), and Chris Kucharik (Director, Wisconsin Environmental Mesonet / Wisconet). Following the panel, UW students from Ankur Desai’s meteorological measurements course will demonstrate some of the standard measurements used in meteorology, including those related to Wisconet.
The Center for Climatic Research focuses on interdisciplinary research regarding the Earth’s climate system, including the causes and impacts of historical, ongoing, and projected climate variability and change, with geographic focuses that span from local to global scale. The center’s diverse activities and societal contributions include the training of students and postdoctoral researchers as the next generation of climate scientists, development of guidance for state government and regional stakeholders regarding climate change impacts and adaptation, and advancement of scientific understanding of the coupled Earth system.
Tools for Tracking Climate Change
CCR scientists have developed easy-to-understand maps that show historical information and climate projections in Wisconsin and Midwest. The maps provide insights into how temperature, precipitation, length of growing season, and other variables might change in the future.
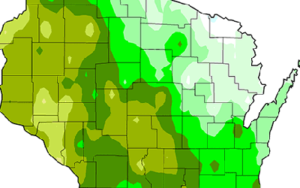
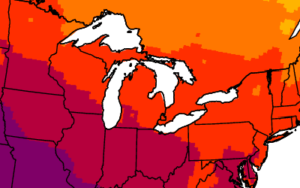
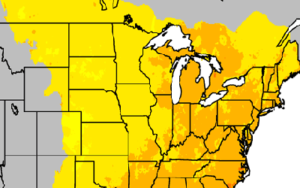
Excellence in Climate Science
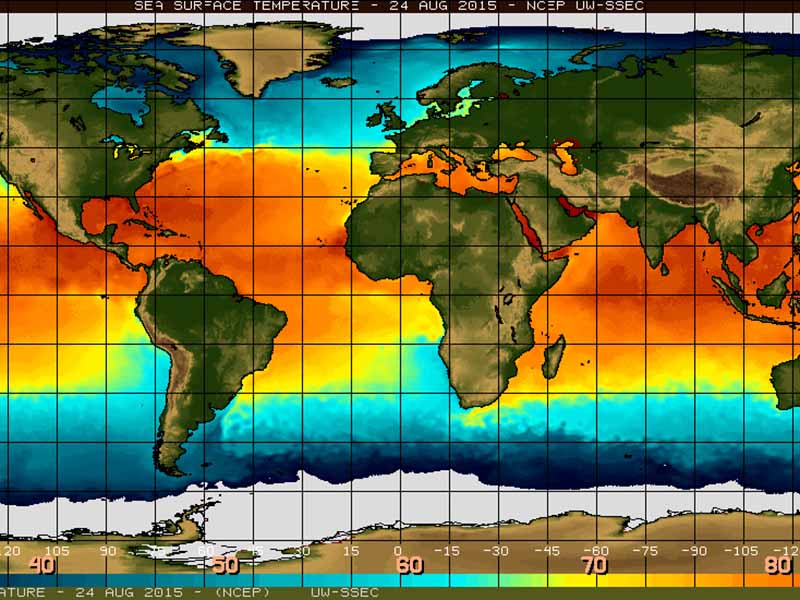
CCR scientists have strong expertise in the areas of climate modeling and climate dynamics, climate impacts, past climates, carbon cycling, terrestrial and aquatic ecosystems, and feedbacks between the atmosphere and earth’s surface.
CCR has a distinguished tradition of over 50 years of excellence in climate science. Its past directors include fellows of the American Geophysical Union and members of the National Academy of Sciences, while its alumni are found in leading atmospheric and geoscience departments around the country. CCR faculty and scientists are at the leading edge of their disciplines.

News and Events
Latest News
- Jack Williams Shares His Opinion With Newsweekon April 11, 2024
- UW–Madison Earth Fest offers education, inspiration, motivation with nearly 50 campus eventson April 9, 2024
- Are we breaking the Atlantic Ocean?on March 18, 2024
- CCR’s Nikaan Koupaei-Abyazani was one of 14 UW-Madison Fulbright award winnerson February 13, 2024
- CCR affiliate, Hannah Zanowski, was awarded the UW Provost’s Exceptional Service Awardon January 30, 2024
Upcoming CPEP Seminars
Missing Ice and altered Ecosystems: Climate Lessons from a Warmer (Early Holocene) Greenland
Supporting the Vitality of Rural Communities through the Wisconsin Rural Partnerships Institute
Building Climate Resilience for Wisconsin Lakes
Support CCR
The Center for Climatic Research advances understanding of the climate system through interdisciplinary investigations of past, present, and future climates and the application of this knowledge for societally relevant purposes. Gifts to CCR play a crucial role in enhancing the breadth and quality of our research and educational programs.